



В. В. Мякотных
О трансформации мышечных волокон в процессе спортивной тренировки / В.В. Мякотных // Вестник спортивной науки. — 2019. — №2. — С. 14 — 20.
Введение. Бурное развитие физиологии спорта, наблюдающееся в последнее время, привело к реальной возможности построения теории физической подготовки спортсменов на основе использования концептуального и математического моделирования. Одним из практических примеров такого подхода может служить разработанная В.Н. Селуяновым концепция протекания специфических приспособительных процессов энергетического метаболизма в мышечных тканях при работе различной направленности [1, 2]. Однако многие специалисты, взявшие на вооружение предложенные автором практические рекомендации, обнаружили низкую эффективность, а иногда и несостоятельность таких занятий. Специальные сайты в интернете пестрят многочисленными вопросами спортсменов и тренеров, не находящими ответов в рамках предложенной концепции. По отдельным сведениям, дающим представление об особенностях подготовки ведущих спортсменов России, создается впечатление, что кризис в некоторых циклических видах спорта, наблюдавшийся в последнее время, во многом связан с попытками перенести положения предложенной теории в практику спортивной тренировки сборных команд страны.
Вот основные положения концепции В.Н. Селуянова [1], не находящие подтверждения при аналитической обработке результатов современных исследований:
1. В структуре мышечной ткани различают медленно сокращающиеся (МС) и быстро сокращающиеся (БС) мышечные волокна (МВ), классифицируемые по содержанию и активности фермента мышечного сокращения АТФазы. Эта мышечная композиция наследуется. В БС-волокнах высока активность АТФазы по сравнению с МС-волокнами, что определяет быструю скорость их мышечного сокращения. В процессе тренировки практически нельзя существенно менять врожденную АТФазную активность MB.
2. Существует классификация MB по содержанию в них митохондрий и активности оксидативных энзимов. В этом случае говорят об окислительных, промежуточных и гликолитических MB (ОМВ, ПМВ и ГМВ, соответственно). Масса митохондрий в ГМВ мала, поскольку необходима только для жизни этих клеток в покое, поэтому они легко утомляются. В тренировочном процессе необходимо стремиться к увеличению массы митохондрий в МВ. При этом можно значительно повысить выносливость без потери скорости и силы мышечных сокращений (за счет сохранения высокой активности АТФазы).
3. Максимальная степень аэробной подготовленности достигается в том случае, когда все миофибриллы оплетаются митохондриальной системой так, что образование новых митохондриальных структур становится невозможным. Необходимо применять статодинамические упражнения для гиперплазии миофибрилл в ММВ, что создаст структурную основу образования новых митохондрий.
4. При выполнении физических упражнений мышечные волокна разных типов сокращаются с разной скоростью.
В связи с обнаруженными противоречиями, требующими дальнейшего анализа, целью нашего исследования явилось выявление основных функциональных механизмов перестройки мышечных волокон под воздействием физических тренировок различной направленности.
Для достижения поставленной цели был проведен аналитический анализ более 20 научных статей и исследований отечественных и зарубежных авторов, включая самые последние публикации, посвященные исследованию механизмов физиологических и биохимических перестроек в скелетной мышце в ответ на изменение функциональных требований.
На основе анализа полученных сведений были выявлены основные направления перестройки мышечной композиции, к которым необходимо стремиться в процессе специальной физической подготовки спортсмена.
Результаты исследования и их обсуждение. Первоначальная классификация МВ основывалась на скорости мышечного сокращения [3]. По этому признаку МВ делились на «медленные» (I тип) и «быстрые» (II тип). Было выявлено, что отличительным морфологическим признаком «медленных» волокон является красный окрас, ввиду разветвленной сети капилляров и большого содержания миоглобина [3]. Этим объяснялась высокая оксидативная емкость таких волокон по сравнению с белыми — «быстрыми». Однако, при дальнейших углубленных исследованиях оказалось, что большинство волокон представляют из себя множество вариантов различных оттенков серого цвета, и каждый ученый был вынужден произвольно выбирать, какие серые волокна называть «белыми», а какие «красными». При этом в широком диапазоне от медленного к быстрому менялась и сократимость МВ [4, 5]. В дальнейшем множество вариантов таких волокон стали называть «промежуточными» или IIа типа (быстрые окислительно-гликолитические), в отличие от «чистых» быстрых гликолитических волокон IIb типа [1, 2, 6]. Гистологический анализ показал высокую степень корреляции между активностью миозиновой АТФазы и скоростью мышечного сокращения, поэтому тип мышечного волокна стали определять по активности этого фермента. Однако, активность АТФазы является пластичным, существенно меняющимся при воздействии соответствующих манипуляций показателем [4, 5, 6]. Ресурсы любой биологической системы весьма ограничены, и она никогда не будет поддерживать функцию с избыточными возможностями. Установлено, что длительная физическая тренировка определенной направленности может приводить к перестройке (снижению или увеличению) энергетического метаболизма мышц и переходу МВ из одного типа в другой, при этом параллельно меняется и миозиновая АТФазная активность. Наиболее распространенным является переход из типа IIb в IIа и наоборот [6]. Меньше доказательств в поддержку возможности преобразования в результате тренировки волокон типа I в тип II. Изменчивость АТФазы проявляется не только в процессе долговременной адаптации, но и в зависимости от величины сопротивления двигательному действию. В сокращающемся против нагрузки МВ цикл АТФазы ингибируется нагрузкой, и для различных мышц уровень активности АТФазы падает примерно до трети своего значения по сравнению с условиями без нагрузки [5].
Таким образом опровергается главный постулат концепции В.Н. Селуянова — неизменность врожденной активности миозиновой АТФазы, якобы определяющей скорость мышечного сокращения.
В последнее время способность к скорости и силе сокращения МВ стали определять по преобладающей изоформе тяжелых цепей миозина мышечного волокна (ТЦМ) [4, 5, 7]. Миозиновый фенотип весьма консервативен, однако при определенных воздействиях удается существенно изменить экспрессию (преобразование информации от миозиновых генов в синтез необходимого белка) и обеспечить трансформацию «медленных» волокон в «быстрые» и наоборот [4, 5, 7, 8]. Считается, что ведущую роль в изменении миозинового фенотипа в сторону трансформации сократительной активности мышц играет перемена характера импульсации мотонейронов, задаваемой величиной нагрузки [7]. По современным представлениям мотонейрон, управляя волокнами с помощью частоты импульсации (10 Гц для «медленных» и 50–60 Гц для «быстрых» ДЕ) влияет на экспрессию миозиновых генов, т.е. на миозиновый фенотип МВ [8]. Так, длительная низкочастотная электростимуляция приводила к появлению около 30–40% волокон медленного типа в преимущественно «быстрых» МВ [8].
Определение функциональных свойств по преобладающей изоформе ТЦМ значительно расширило классифицированные МВ. К примеру, в одной из классификаций определяют четыре изоформы и, соответственно, четыре типа волокон: I – «медленный»; IIа – «быстрый»; IId/x – «быстрый»; IIв — «самый быстрый» [7]. В другой признано, что скелетные мышцы человека состоят из трех «чистых» (I, IIа, IIx) и трех гибридных (I / IIa, IIa / IIx, и I / IIa / IIx) типов [4]. Гибриды представляют собой единичные волокна, в которых представлены два или три типа волокон одновременно и изначально предназначены для целесообразной перестройки при адаптации мышц к физической нагрузке определенной направленности. Иногда выделяют волокна IIс типа, проявляющие как окислительную, так и гликолитическую активность и т.п. Возникшая путаница в определении близких оттенков «красных» МВ, а также изоформ ТЦМ, привела к тому, что некоторые специалисты стали вообще утверждать о невозможности классификации МВ ввиду динамичности и гибкости их трансформации.
Широкий диапазон пластичности в ответ на изменение функциональных требований, выражаемый в разнообразии физиологических и биохимических приспособлений в скелетной мышце, ставит вопрос о выборе базового, ключевого показателя, определяющего функциональные свойства мышц. За исходный признак, определяющий все остальные, следует принять показатели мощности той или иной системы энергообеспечения мышечного сокращения, которые имеют относительно четко определенные границы своих возможностей. Нет врожденно неизменных «быстрых» или «медленных» волокон, есть МВ с наследственно детерминированным механизмом энергообеспечения. А тот или иной механизм энергообеспечения определяет потенциальные возможности мобилизации энергетических субстратов, с соответствующим распадом АТФ и выделением свободной энергии для мышечного сокращения. Именно изменение мощности освобождения энергии при перемене характера энергообеспечения и определяет все последующие адаптационные перестройки, обеспечивающие необходимое сочетание скорости, силы сокращения и выносливости МВ в различных видах спорта.
Алактатная система энергообеспечения отличается минимальной сложностью реакции ресинтеза АТФ, быстрой доступностью и мобилизуемостью макроэргов. Поэтому у спортсменов высокого класса мощность алактатной системы может достигать 4,3 кДж/кг/мин [6]. Однако, ограниченная емкость приводит к исчерпанию возможностей этой системы уже через десять секунд после начала работы. В лактатной системе ресинтез АТФ происходит за счет процессов анаэробного гликолиза, сопровождающихся накоплением лактата и ионов Н+ и быстрым развитием утомления. Он осуществляется серией сложных последовательных реакций, поэтому энергетические субстраты менее мобилизуемы, а мощность освобождения энергии уступает алактатной системе и не превышает 3,1 кДж/кг/мин [6]. По сравнению с мощностью алактатной и лактатной анаэробных систем, возможности аэробной системы невелики, и у спортсменов высокого класса составляют 1,5—1,8 кДж/кг/мин [6]. В этом случае мощность лимитируется, в первую очередь, ограниченным потенциалом кислородтранспортной системы, с соответствующим снижением мобилизационных возможностей вовлечения в процесс энергетических субстратов.
В этой схеме скорость сокращения становится производной от мощности системы энергообеспечения в волокнах различного типа. Гликолитические МВ быстро утомляемы, но способны выполнять максимальное количество работы в единицу времени. В соответствие с законом Хилла, при незначительном внешнем сопротивлении движению такие ГМВ становятся быстрыми, при выраженном внешнем сопротивлении — медленными (сопротивление ингибирует цикл миозиновой АТФазы), но сильными (за счет образования множества актомиозиновых «мостиков»). Мало утомляемые ОМВ, ввиду использования аэробных механизмов энергообеспечения способны выполнять меньшее количество работы в единицу времени, поэтому потенциально более медленные. В большинстве скоростно-силовых упражнений оба типа таких МВ могут сокращаться с одинаковой скоростью, но разным вкладом в обеспечение движения. Реальная скорость мышечных сокращений в скоростно-силовых упражнениях значительно ниже потенциальной (без нагрузки). При этом «мощные» гликолитические МВ, хотя и не реализуют полностью свои скоростные возможности, но вносят основной вклад в обеспечение «тяги» мышц, а «маломощные» окислительные МВ тратят практически весь свой энергопотенциал на обеспечение скорости сокращения, при минимальном вкладе в тяговые характеристики. В этом случае изменение активности АТФазы в зависимости от величины нагрузки и играет роль «синхронизатора» скорости сокращений МВ различных типов. И, наконец, существует множество вариантов промежуточных МВ с обратно пропорциональным соотношением между мощностью и длительностью работы, обусловленным разным сочетанием анаэробных и аэробных энергетических процессов (повысить выносливость ГМВ возможно только за счет частичного замещения гликолиза тканевым дыханием, т.е. за счет потери мощности, и наоборот). Это сочетание и определяет оптимальную мышечную композицию для различных видов спорта, к формированию которой необходимо стремиться в процессе специальной физической подготовки спортсмена.
Считается, что наиболее полное представление о процессах долговременной адаптации скелетных мышц к упражнениям различной направленности дает моделирование параметров нагрузки с помощью хронической импульсации различной частоты [5, 8, 11]. Наиболее показательные эффекты перестройки фенотипа скелетной мышцы достигались путем длительной низкочастотной стимуляции в 10 Гц, наложенной на преимущественно «быструю» мышцу, что моделировало нагрузку умеренной интенсивности. При этом наблюдались четко выраженные прогрессирующие изменения, в процессе которых «быстрая» мышца сначала меняла свой энергетический метаболизм, затем свои сократительные свойства, полностью трансформируясь в «медленную» с соответствующим «глубоко-красным» окрасом.
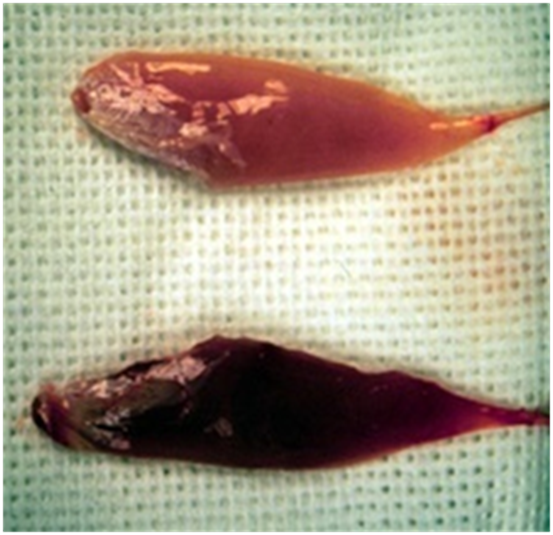 Рис. 1. Фотография контрольной (верхней) и экспериментально стимулированной (нижней) мышцы голени кролика.
Рис. 1. Фотография контрольной (верхней) и экспериментально стимулированной (нижней) мышцы голени кролика.
Хронология изменений имела следующую последовательность [11]:
1. 3 часа после начала стимуляции. Увеличение в объеме и объединение сети мембран саркоплазматического ретикулума. Значение этого морфологического изменения пока не выяснено.
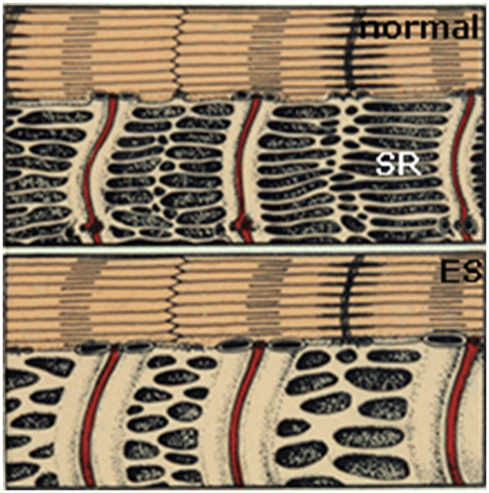 Рис. 2. «Распухание» сети мембран саркоплазматического ретикулума через несколько часов после начала стимулирования.
Рис. 2. «Распухание» сети мембран саркоплазматического ретикулума через несколько часов после начала стимулирования.
2. От 2-х до 12 дней стимуляции. Гиперплазия митохондриальной системы, повышение активности окислительных ферментов и плотности капилляров. Значение этих изменений заключается в перестройке энергетического метаболизма к длительной работе умеренной интенсивности за счет увеличения окислительной способности МВ. При этом запускается экспрессия ряда факторов транскрипции, активирующая гены ядерного и митохондриального генома, необходимые для синтеза митохондрий [13, 14].
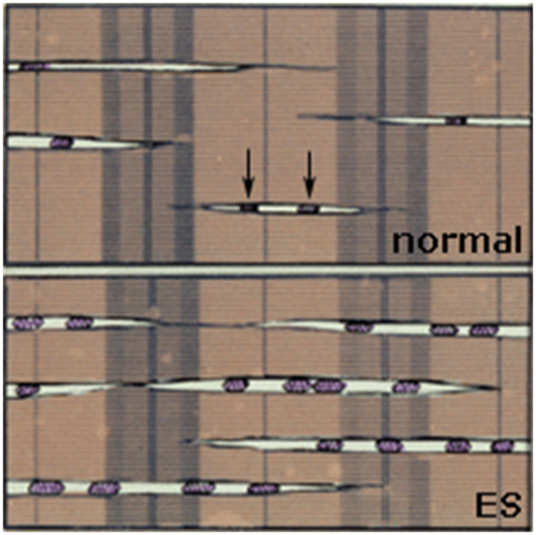
Рис. 3. Увеличение размера и количества митохондрий (обозначенных стрелками до стимуляции), присутствующих в ткани в результате хронической стимуляции.
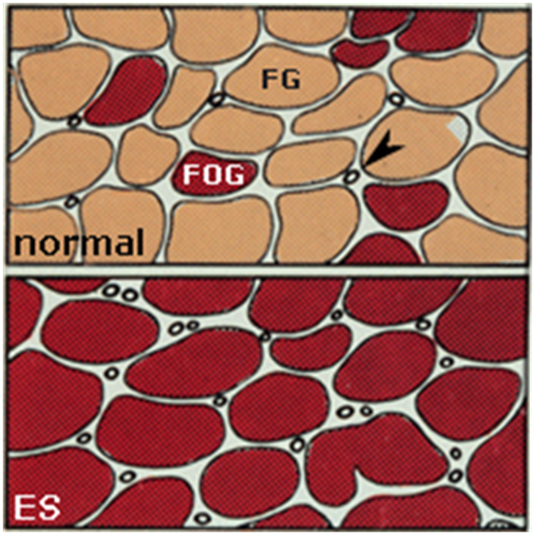 Рис. 4. Гистохимические изменения в виде появления волокон с «глубоко красным» окрасом и увеличения количества капилляров (обозначены стрелкой).
Рис. 4. Гистохимические изменения в виде появления волокон с «глубоко красным» окрасом и увеличения количества капилляров (обозначены стрелкой).
3. 14 дней стимуляции. Увеличение ширины z-диапазона МВ, снижение содержания и активности Са2+-АТФазы. Снижение экспрессии количества и активности транспортных белков кальция саркоплазматического ретикулума физиологически объясняется необходимостью создания структуры, соответствующей новым функциональным требованиям, связанным с увеличением времени выхода на пик напряжения и периода релаксации МВ.
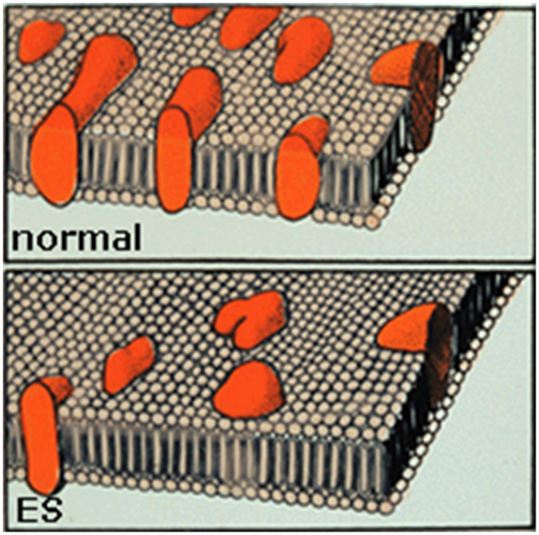 Рис. 5. Уменьшение количества, активности и изменение распределения Са2+-АТФазы.
Рис. 5. Уменьшение количества, активности и изменение распределения Са2+-АТФазы.
4. 28 дней стимуляции. Изменение профиля световой цепи миозина, при котором в нормально быстрой мышце, содержащей только легкие цепи LC1f, LC2f и LC3f, появлялись легкие цепи, характерные для «медленных» волокон (LC1s и LC2s)
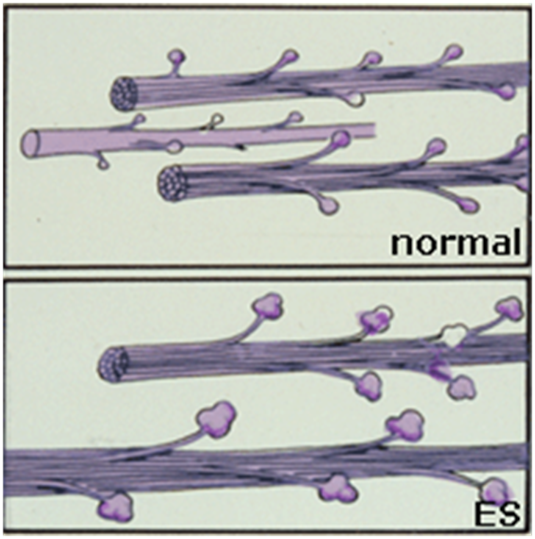 Рис. 6. Изменение профиля световой цепи миозина, при котором в нормально быстрой мышце появляются легкие цепи, характерные для медленных волокон.
Рис. 6. Изменение профиля световой цепи миозина, при котором в нормально быстрой мышце появляются легкие цепи, характерные для медленных волокон.
5. 28 дней стимуляции. Снижение массы скелетных мышц и площади поперечного сечения волокон. Объясняется физиологической необходимостью уменьшения диффузионных расстояний от мышечного волокна до промежуточных пространств, содержащих капилляры.
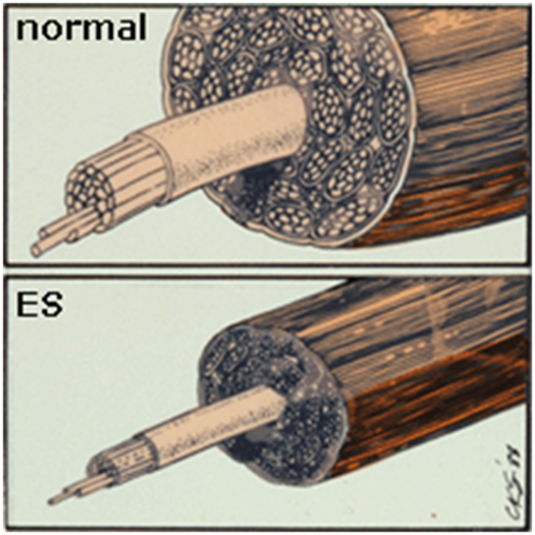 Рис. 7. Значительное уменьшение площади поперечного сечения и мышечной массы мышечных волокон.
Рис. 7. Значительное уменьшение площади поперечного сечения и мышечной массы мышечных волокон.
С практической точки зрения выявляется спорность идеи В.Н. Селуянова о необходимости применения статодинамических упражнений для гипертрофии миофибрилл окислительных МВ, создающей условия образования новых митохондрий. Наблюдаемое в эксперименте увеличение размеров и количества митохондрий выглядит далеко не критическим для применения таких манипуляций (рис.3). К тому же такая гипертрофия вступает в явное противоречие с наблюдаемым природным механизмом адаптации, связанным с уменьшением массы и площади поперечного сечения МВ, а значит с облегчением доставки кислорода к митохондриям окислительных МВ (рис. 6). Любая заметная гипертрофия миофибрилл окислительных МВ свидетельствует о включении анаэробного гликолиза в процесс энергообеспечения, с соответствующей перестройкой миозинового фенотипа на большую мощность мышечных сокращений. Очевидно дальнейшее совершенствование системы тканевого дыхания лежит не в гиперплазии митохондриальной системы, а в развитии сети капилляров работающих мышц, повышении активности окислительных ферментов и эффективности работы митохондрий, формировании механизмов их защиты от окислительного стресса, развития способности к удалению и утилизации лактата в промежуточных МВ. Силовая тренировка, как база обеспечения оптимальных для данного вида спорта силовых способностей, повышения содержания мышечного гликогена, клеточных органелл (в т.ч. и митохондрий), возможна только на относительно небольшом отрезке общеподготовительного этапа подготовительного периода.
Результаты моделирования показывают, что даже «чисто» аэробная циклическая работа, с интенсивностью ниже порога рекрутирования ПМВ, не только увеличивает окислительный потенциал «медленных» МВ, но и приводит к энергетической перестройке высокопороговых ДЕ, значительно повышая их аэробные возможности. По В.Н. Селуянову такое возможно лишь при нагрузке, соответствующей достижению порога рекрутирования этих волокон, поэтому отвергается объемная базовая работа умеренной интенсивности. Становится понятным физиологический смысл развития «общей выносливости» на базовом этапе подготовительного периода. Увеличение количества ОМВ включенных в работу, возрастание их энергопотенциала за счет «неутомляемых» аэробных систем энергообеспечения, позволяет перевести часть энергоресурсов этих систем на повышение скоростных характеристик движения при сохранении должных показателей «тяги».
Мощность аэробной энергосистемы строго ограничена возможностями этой системы, и обеспечивает хороший результат только в циклических видах спорта с работой умеренной интенсивности (сверхдлинные дистанции в лыжных гонках, беге, плавании, спортивной ходьбе). Переход в зону интенсивности, превышающую возможности аэробного метаболизма, возможен только при подключении более мощных гликолитических механизмов, за счет повышения экспрессии генов белков анаэробного гликолиза и индуцирования процессов митофагии (разрушения митохондрий). Причем чем больше вес спортсмена, тем мощнее мышечные напряжения, необходимые для обеспечения движения, тем выше вклад в работу гликолиза с соответствующим закислением и развитием утомления.
Несмотря на недостаточную изученность механизмов частичного замещения тканевого дыхания гликолизом, можно с уверенностью утверждать, что экспрессия факторов гликолиза до значений, превышающих возможности буферных систем, приводит к снижению экспрессии факторов аэробного метаболизма и наоборот [16]. Один из механизмов можно представить следующим образом: активация гликолиза вызывает выход ионов Н+ в саркоплазму и закисление, приводящее к высвобождению ионов Fe2+ из комплекса с трансферрином [18]. Каждая молекула трансферрина при нейтральной кислотности среды присоединяет два иона железа. Однако при закислении способность трансферина связывать железо снижается, и оно легко высвобождается из ставшего непрочным комплекса. При этом свободное железо обладает способностью стимулировать образование гидроксил-радикала, т.е. повышать в клетке содержание активных форм кислорода (АФК). Близость к дыхательной цепи и отсутствие защитных гистонов вызывает повреждения митохондриальной ДНК, изменяет дыхательную цепь, вторично увеличивая генерацию свободных радикалов. В ответ на дисфункцию окислительной системы фосфорилирования клетки активируют «митохондриальный развернутый белковый ответ» (mtUPR) для восстановления митохондриальной функции. Если, несмотря на активацию mtUPR, производство АФК не снижается, митохондрии направляются на путь митофагии (самоуничтожения) [19, 20].
В этой тонкой, постепенной настройке мышечной композиции, приближающей сочетание аэробных и анаэробных путей освобождения энергии к оптимальному для данного вида спорта (на фоне дальнейшего совершенствования механизмов тканевого дыхания и гликолиза) и заключается физиологический смысл специально-подготовительного этапа подготовки спортсмена. Сначала речь идет о постепенном приближении к соревновательной скорости непрерывной «темповой» работой на грани ПАНО, при сохранении запланированного объема. Затем в процесс включается интервальная тренировка, причем скорость (интенсивность) на отрезках дистанции должна приближаться к запланированной на первых стартах, но не превышать ее. Следует учесть, что адаптация развивается строго специфично к характеру предъявляемых требований. Даже одна высокоинтенсивная и объемная тренировка, неспецифичная для предстоящей соревновательной деятельности и превышающая текущие биологические возможности спортсмена, может привести к массовой митофагии и перестройкам, сводящим на нет весь, накопленный в предыдущий период, потенциал аэробного энергообеспечения.
В последнее время выясняется, что включение гликолиза в процесс энергообразования запускает мощные приспособительные процессы защиты митохондриального генома от окислительного стресса. Адаптационные перестройки в этом направлении особенно важны в видах спорта, с нагрузками субмаксимальной интенсивности, а также с ситуативной нагрузкой, сочетающей фазы работы максимальной и умеренной мощности. Вероятно, в этом случае приспособительные реакции развиваются по следующим основным направлениям:
1. Перестройка метаболизма в митохондриях. Экспрессия генов белков, выводящих лактат для «сжигания» в митохондрии, и через мембрану мышечной клетки в сосудистое русло. Тем самым удается значительно снизить локальное закисление работающих мышц. Установлено, что в процессе адаптации к интенсивной тренировке содержание белков такого «транспортного насоса» в клетке может увеличиваться на 70% [20].
2. Активизация механизмов защиты митохондрий от окислительного стресса. Считается, что высокоинтенсивная физическая нагрузка имеет отрицательное влияние на антиоксидантную систему [21]. С другой стороны, установлено, что у организмов с интенсивным производством АФК происходит изменение генетического кода в системе биосинтеза белка в митохондриях — кодон AUA начинает кодировать метионин вместо изолейцина. Содержание поверхностных метионинов в митохондриальном цитохроме может увеличиваться в 10 раз. Метионин обладает антиоксидантной активностью и способен реагировать практически со всеми природными АФК, включая даже относительно инертную перекись водорода. При этом АФК обезвреживаются, а метионин превращается в стабильный метионинсульфоксид [22].
3. Восстановление функции митохондрий за счет повышения экспрессии генов, кодирующих белки «митохондриального развернутого белкового ответа» [19].
Приспособительные процессы в этих направлениях развиваются постепенно и параллельно с формированием оптимальной мышечной композиции. Любое неоправданное форсирование интенсивности нагрузок, при ещё незавершенных настройках защиты от окислительного стресса, может привести к повреждениям митохондриального генома и снижению эффективности тканевого дыхания. Именно в разработке упражнений, целенаправленно снижающих степень локального закисления МВ и формирующих механизмы защиты от окислительного стресса для различных видов спорта и кроется потенциал повышения эффективности развития специальной выносливости в тренировке.
Выводы:
1. Успех в современной системе спортивной тренировки во многом определяется эффективностью генетических перестроек врожденной мышечной композиции. Именно количество МВ определенного типа в значительной мере обусловливает достижения спортсменов в том или ином виде спорта.
2. Мышечная композиция генетически детерминирована, однако под воздействием тренировок определенной направленности происходит существенное изменение экспрессии генов и, соответственно, морфологических, физиологических и биохимических свойств МВ.
3. Пластичность МВ выражается в перестройке энергетического метаболизма, изменении сократительной способности мышц и активности ферментов мышечного сокращения, трансформации биосинтеза белка в митохондриях для защиты от окислительного стресса.
4. Трансформация свойств МВ различных типов начинается с перестройки систем энергообеспечения, в сторону формирования оптимального, для предъявляемой нагрузки, сочетания мощности и выносливости мышечных сокращений. Это ключевой фактор, определяющий все последующие изменения.
5. Чем значительнее «идеальная» модель мышечной энергетики отличается от генетически детерминированной, тем больше усилий необходимо затратить на перестройку, тем слабее закрепленные изменения и быстрее процессы реадаптации при снижении специфических требований. Неудачный врожденный фенотип МВ может свести к минимуму эффективность самой современной системы спортивной тренировки.
Сокращения:
АДФ — аденозиндифосфат
АФК — активные формы кислорода
ДЕ — двигательная единица
МВ — мышечные волокна
ГМВ — гликолитические мышечные волокна
ОМВ — окислительные мышечные волокна
ПМВ — промежуточные мышечные волокна
ПАНО — порог анаэробного обмена
ТЦМ — тяжелые цепи миозина
mtUPR — митохондриальный развернутый белковый ответ
1. Селуянов, В.Н. Спортивная адаптология / В.Н. Селуянов. — В кн. «Энциклопедия систем жизнеобеспечения». — Магистр пресс/Юнеско. -2011. — 1000 с.
2. Селуянов, В.Н. Эмпирический и теоретический пути развития теории спортивной тренировки / В.Н. Селуянов // Теория и практика физической культуры. — 1998. — №3.- С. 46-50.
3. Sheep, M. of The fast and slow muscles of the rabbit / M. Sheep, K. Sheep, T.Rekard // ArchBiochemBiophys. — 1965. — №109.-Р.185-191.
4.Biral, D. Сomposition of myosin heavy chain of individual fibers of normal human muscle / D. Biral, R. Betto, D. Danieli-Betto, G.Salviati //Biochem J. — 1988. — 250 (1). — Р. 307-308.
5.Walklet,J. Isoforms of myosin and cross-mechanochemical cycle / J. Walklet, Z. Ujfalusi, M. A. // Journal of experimental biology. -2016. -№219: Р. 168-174; doi: 10.1242 / jeb.124594
6. Платонов,В.Н. Система подготовки спортсменов в олимпийском спорте. Общая теория и ее практические приложения / В.К. Платонов. — К.: Олимпийская литература. — 2004. — 808 с.
7. Шенкман, Б.С. От медленных к быстрым. Гипогравитационная перестройка миозинового фенотипа мышечных волокон / Б. С. Шенкман. — ACTA NATURAE. — 2016. — Том 8. — № 4 (31). — С. 52-65
8. Pette, D. Skeletal muscle plasticity. In: Skeletal muscle plasticity in health and disease / D. Pette // C. Springer. — 2006. P. 1–27.
9. Ланская, О.В. Изучение уровня возбудимости кортикоспинальных и нервно-мышечных структур у представителей различных видов спорта / О.В. Ланская, Е.В. Ланская, Е.Ю. Андриянова //Ульяновский медико-биологический журнал. — 2015. — № 3. — С. 101 — 112.
10. Андриянова, Е.Ю. Физиологические механизмы функциональной пластичности спинальных систем двигательного контроля при занятиях спортом / Е.Ю. Андриянова, О.В. Ланская. — Великие Луки : Великолукская государственная академия физической культуры и спорта. — 2013. — 231 c.
11. Lieber, R.L. Time and cellular control of muscle fiber type transformation after chronic stimulation /R.L. Lieber // ISI Atlas of Science. -1988: 1. — Р.189-194.
12. Yan, Z. Regulation of exercise-induced fiber type transformation, mitochondrial biogenesis and angiogenesis in skeletal muscles / Z. Yan, M.Okutsu, Y. N. Akhtar, V. A. Lira // Journal of Applied Physiology . 2011. — V. 110/ — P. 264-274.
13. Yan, Z. Regulation of mitochondrial quality under the influence of exercise/ Z. Yan, V. A. Lira, N.P.// Exerc Sport Sci Rev. — 2012. -V. 40 (3). -P.159-164
15. Baker, J. S. Interaction among Skeletal Muscle Metabolic Energy Systems during Intense Exercise / J. S. Baker, M. C. McCormick, R. A. Robergs // Journal of Nutrition and Metabolism. — 2010, ID 905612, 13 pages. — http://dx.doi.org/10.1155/2010/905612
16. Moyes, C. D. Control of muscle bioenergetic gene expression: effect of glycolytic and oxidative enzymes on allometric scaling / C. D. Moyes, C. M. R. Lemoine // Journal Of Experimental Biology. — 2005. — V. 208. — P.1601-1610.
17. Abe, T. High-intensity interval training-induced metabolic adaptation combined with increased HIF-1α and glycolytic protein expression / T. Abe, Y. Kitaoka, D.M.Kikuchi, K.Takeda , O. Numata // J Appl Physiol. — 2015/ — V. 119 (11). — P. 1297-1302.
18. Moreira, O. C. Mitochondrial function and mitophagy in the elderly: effects of exercise / O. C. Moreira, B. Estébanez, S. Martínez-Florez, J. Paz, M. J. Cuevas, J. González-Gallego // Oxidative medicine and cellular longevity Vol.2017. — ID 2012798, 13 pages. — https://doi.org/10.1155/2017/2012.79
19.Tian, Y. “Mitochondrial UPR: a double-edged sword” / Y. Tian, C. Merkwirth, A. Dillin // Trends in cell biology. -2016. — Vol.26 (8). -https://doi.org/10.1016/j.tcb.2016.06.006
20.Juel, C. The effect of high-intensity periodic training on lactate and H+ release from human skeletal muscle / C. Juel, C. Klarskov, J. J. Nielsen, P. Krustrup, M. Mohr, J. Bangsbo // American Journal of Phisiologi. — 2004. — Vol. 286(2). — P. 245-251. — https://doi.org/10.1152/ajpendo.00303.2003
21. Мякотных, В.В. Двигательная активность и возрастная инволюция функциональных возможностей человека / В.В. Мякотных. – Сочи: СГУ, 2013. – 176 с.
22. Скулачев, В.П. Новые сведения о биохимическом механизме запрограммированного старения организма и антиоксидантной защите митохондрий / В. П. Скулачев // Биохимия. — 2009. — Том 74. — Вып. 12. — С. 1718 — 1721.
23. Мякотных, В.В. О трансформации мышечных волокон в процессе спортивной тренировки / В.В. Мякотных // Вестник спортивной науки. — 2019. — №2. — С. 14 — 20.